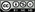Corteza prefrontal e insula en el procesamiento emocional
Si bien era asumida desde antiguo la participación de la corteza cerebral en el procesamiento emocional, no fue hasta los años 20 cuando Cannon comenzó a realizar los primeros experimentos científicos a este respecto, estableciendo el papel del tálamo como suministrador general de información sensorial a la corteza. El procesamiento emocional quedaba entonces dividido en una respuesta periférica, coordinada por el hipotálamo, y un procesamiento cortical a partir de la aferencia talámica, aunque la relación entre ambos estadios aún no se conocía.
Unos años más tarde, James Papez propuso que el mecanismo cortical encargado del procesamiento de los "sentimientos" (por aquel entonces se reservaba el término "emoción" para la manifestación somática) quedaba contenido en lo que Broca había denominado lóbulo límbico, y que comprendía la circunvolución del cuerpo calloso (cíngulo), la circunvolución parahipocámpica (prolongación ántero-inferior del cíngulo), y la formación hipocámpica (hipocampo, giro dentado y subículo). Este autor describió la manera en que la respuesta somática y el procesamiento emocional cortical se relacionaban anatómicamente, a través de dos sistemas complementarios, en lo que desde entonces se conoce como el circuito de Papez: la información aportada por el tálamo al lóbulo límbico se proyecta al hipocampo, que pondrá en comunicación esta información con el hipotálamo y el área septal a través del fórnix (sistema hipocampo-septal). Desde los cuerpos mamilares del hipotálamo, a su vez, la información volvería a los núcleos anteriores del tálamo por medio del haz mamilo-talámico (sistema mamilo-talámico), desde donde se proyectaría de vuelta a la corteza. La observación de que pacientes infectados por el virus de la rabia, que afecta de forma característica el hipocampo, padecían severas alteraciones en su estado emocional (manifestando accesos de temor y rabia), parecía dar validez a esta teoría, aunque Papez sobredimensionó el papel del hipocampo en el procesamiento emocional (papel de coordinación que es ejercido en su lugar por la amígdala).
MacLean amplió posteriormente los límites del lóbulo límbico, añadiendo como partes implicadas en el procesamiento emocional la corteza orbitofrontal, el área septal, el núcleo accumbens, y más notoriamente la amígdala. Completan finalmente los órganos del sistema límbico la corteza de la ínsula y el polo temporal. Mesulam añade un elemento anatómico y citoarquitectónico a este esquema, agrupando por un lado el polo temporal, la ínsula y la corteza orbitofrontal en una subdivisión "olfatocéntrica" del cinturón paralímbico, como transición del alocórtex olfatorio y amigdalino al isocórtex, y por otro lado el cíngulo y la corteza parahipocampal como subdivisión "hipocampocéntrica", como transición del alocórtex hipocampal nuevamente al isocórtex (Mesulam MM 1998).
En 1939, Klüver y Bucy describieron el resultado de la extirpación temporal bilateral en monos (incluyendo la corteza temporal extra-límbica), en lo que supuso una nueva prueba de la participación del lóbulo límbico en el procesamiento emocional: los animales se amansaban, desarrollaban hiperoralidad e hipersexualidad, tendían compulsivamente a observar y reaccionar ante estímulos visuales, y tenían dificultades para el reconocimiento de objetos familiares. El síndrome de Klüver-Bucy fue explicado en un principio siguiendo la idea neuroanatómica de Papez, pero con el tiempo se pudo comprobar cómo tanto el hipocampo como los cuerpos mamilares y núcleos anteriores del tálamo no participan de una manera sustancial en el procesamiento emocional, sino más bien en procesos de almacenamiento de información (memoria), y por otro lado cómo el déficit visual asociativo dependía más de la lesión de la corteza temporal inferior, que de estructuras pertenecientes al propio lóbulo límbico.
El núcleo ventral posterior parvocelular del tálamo, que recibe la aferencia sensorial visceral, transmite la información a la corteza insular anterior (gusto, plenitud abdominal, punzadas del hambre), donde existe un mapa topográfico del sistema de los órganos internos (Kandel ER 2001): en la porción más anterior está representado el gusto, después las sensaciones gastrointestinales, y más dorsalmente la representación cardio-respiratoria. Esta corteza sensorial visceral (ínsula) interactúa con el extremo anterior de la circunvolución del cuerpo calloso, en un área llamada "infralímbica", que representa una verdadera área motora visceral, cuya estimulación da lugar a las correspondientes respuestas viscerales, mediante su proyección eferente hacia el resto de estructuras del sistema autónomo central. Las lesiones en la corteza sensorial visceral dan lugar a una pérdida en la capacidad de apreciación consciente de sensaciones como el gusto, mientras que las lesiones en la corteza visceral motora se relacionan con la abulia, estado caracterizado por una pérdida de la manifestación de las reacciones emocionales que normalmente acompañan a los estímulos externos.
La corteza orbitofrontal está implicada en la ordenación de la conducta en base al valor de recompensa o castigo de los estímulos ambientales, en un sistema de vigilancia frontal que busca adecuar los impulsos a la realidad, de tal manera que su lesión da lugar a desinhibición social, impulsividad y conductas inapropiadas, así como a malinterpretación de expresiones emocionales en otros sujetos (componente de teoría de la mente y cognición social). Se ha visto así cómo las neuronas de la corteza orbitofrontal responden por ejemplo a la recompensa o el castigo monetarios, y lo hacen además de una manera proporcional a la magnitud de dicho refuerzo (Hare TA 2008). También se ha relacionado la disfunción de esta región cortical con que los sujetos se vuelvan más sensibles a la recompensa pasada, y a la vez menos sensibles al castigo (o incluso a nuevas recompensas), lo que condiciona el desarrollo de síntomas de impulsividad, irritabilidad y frustración ante una no recompensa (Berlin HA 2004), si bien esta tendencia puede también deberse a una menor evaluación de las posibles consecuencias de los actos, ya sean positivas o negativas, posiblemente por una aceleración en la percepción del tiempo y en el ritmo cognitivo subjetivo (cognitive tempo). Como síntoma genérico de los síndromes prefrontales, también se observa en estos casos una cierta perseverancia en la conducta, más vinculada a lo emocional que a lo ejecutivo, en el sentido de tender el sujeto a seguir respondiendo invariablemente y de manera inflexible a determinados estímulos sensoriales, aun cuando ya no se encuentren vinculados con una recompensa, o se acompañen en el nuevo contexto de un castigo. Estos datos indican en conjunto que la corteza orbitofrontal no actúa simplemente inhibiendo respuestas previas, innatas o adquiridas, sino que participa en una regulación conductual más compleja, como el proceso fisiológico de extinción que ocurre en el aprendizaje normal, o el mecanismo de aprendizaje inverso guiado emocionalmente. Resulta interesante constatar finalmente cómo los sujetos con daño orbitofrontal mantienen una noción consciente y congruente de su déficit, así como una reacción emocional adecuada al fracaso y la frustración que experimentan (frecuentes suspiros, muecas, gestos de desagrado), pero no por ello logran controlar su conducta impulsiva, lo que indica una preservación de los mecanismos explícitos de monitorización de la conducta, contrapuesta al daño en los sistemas implícitos de ajuste de asociaciones entre estímulos y refuerzos cambiantes.
La formación hipocámpica, a pesar de no formar el núcleo principal del sistema límbico (a diferencia de lo sugerido por Papez), sí tiene una participación indirecta en el procesamiento emocional. Como estructura implicada en la formación de memoria explícita (consciente) y en el condicionamiento contextual, participa en el recuerdo de las características explícitas de las circunstancias en las que tuvo lugar el estímulo cargado emocionalmente (LeDoux J 1999), de tal manera que su lesión condiciona la aparición de un déficit en el conocimiento contextual de los estímulos emocionales (cuándo ocurrió, en qué sitio, de qué manera).
La corteza prefrontal ventromedial -al margen del cíngulo y de la corteza orbitofrontal medial- desempeña un papel complejo en la cognición emocional y social, participando en los estados de motivación y en la generación de iniciativa, en la prospección de posibles consecuencias de nuestras acciones y el mantenimiento y monitorización de nuestros planes en el tiempo -memoria prospectiva- (Okuda J 1998), así como en la toma de decisiones (Bechara A 1998), dando lugar su lesión a cuadros de apatía, inhibición, retraimiento social y mutismo acinético. El polo frontal dirige y pondera además los procesos cognitivos, bien en forma de una atención orientada al estímulo (escuchar en una conversación, ver televisión, leer), bien de una atención independiente del estímulo (insight, pensamiento), en coordinación con áreas posteriores de índole gnósica (precuneus), en lo que se ha denominado "gateway hypothesis" (Burgess PW 2007).
Las cortezas orbitofrontal y prefrontal ventromedial reciben una aferencia multimodal procedente de los cinco sentidos, lo que junto con sus numerosas conexiones límbicas -hipótesis del marcador somático- (Damasio A 2006) les permite determinar el valor de recompensa de los estímulos, atribuyendo un significado afectivo a los objetos de la percepción (especialmente a percepciones complejas como la voz o el rostro). El grado de dependencia con respecto a la recompensa conformará patrones de conducta y rasgos de personalidad (preferencias, temperamento), que el aprendizaje social matizará posteriormente (Murphy BL 1996). Esta valoración de las circunstancias externas no sólo conformará nuestra conducta, sino también nuestro propio entendimiento del medio externo, con una participación no sólo en el proceso de la cognición social sino de la misma consciencia de uno mismo (Esposito F 2010, Ebstein RP 2010, Rankin KP 2006).
REFERENCIAS
Murphy BL, Arnsten AF, Goldman-Rakic PS, et al. Increased dopamine turnover in the prefrontal cortex impairs spatial working memory performance in rats and monkeys. PNAS 1996; 93(3): 1325-1329.
Esposito F, Rochat L, Van der Linden A-CJ, et al. Apathy and Executive Dysfunction in Alzheimer Disease. Alzheimer Disease & Associated Disorders 2010; 24(2): 131-137.
Ebstein RP, Israel S, Chew SH, et al. Genetics of Human Social Behavior. Neuron 2010; 65(6): 831-844.
Rankin KP, Gorno-Tempini ML, Allison SC, et al. Structural anatomy of empathy in neurodegenerative disease. Brain 2006; 129(11): 2945-2956.
Damasio A. El error de Descartes. Editorial Crítica. Barcelona, 2006.
Mesulam MM. From sensation to cognition. Brain 1998; 121: 1013-1052.
Berlin HA, Rolls ET, Kischka U. Impulsivity, time perception, emotion and reinforcement sensitivity in patients with orbitofrontal cortex lesions. Brain 2004; 127: 1108-1126.
LeDoux J. El cerebro emocional. Editorial Planeta. Barcelona, 1999.
Okuda J, Fujii T, Yamadori A, et al. Participation of the prefrontal cortices in prospective memory: Evidence from a PET study in humans. Neuroscience Letters 1998; 253: 127-130.
Bechara A, Damasio H, Tranel D, et al. Dissociation of working memory from decision making within the human prefrontal cortex. J Neurosci 1998; 18(1): 428-437.
Burgess PW, Dumontheil I, Gilbert SJ. The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends in Cognitive Sciences 2007; 11 (7): 290-298.
Hare TA, O´Doherty J, Camerer CF, et al. Dissociating the Role of the Orbitofrontal Cortex and the Striatum in the Computation of Goal Values and Prediction Errors. J Neurosci 2008; 28(22): 5623-5630.
- 50060 lecturas